 |
| Image 1: Ever thought why your granny had to take her cod-liver oil, whenever there she suffered from an ailment as a child? Probably not due to the rancid omega-3 oils which were partly responsible for its awful taste. That its high vitamin A content may not just have helped her to ward off the colds and infections, but also to stay lean, is yet a potential "side-effect" of dietary retinol for clear-cut which evidence is emerging only recently. |
Facebook followers of mine have probably seen my post on the beta carotene metabolites called β1-apocarotenoids - recently discovered naturally occurring vitamin A receptor antagonist (block the activity or "real" vitamin A = retinol) about two to three days ago (
Eroglu. 2012). As a
SuppVersity regular, you will also be aware that vitamin A in its active form is
not simply a dangerous substance that is to be avoided at all costs. And though beta carotene has long lost its image as (yet another) super-vitamin, with both direct supplementation and food-enrichment being scrutinized, the aforementioned results yield a couple of interesting hypothesis that may be worth investigating - if we also take into account the results of another recently published study that was conducted by researchers from the
Department of Biophysical Chemistry at
Kyoto Pharmaceutical University in Kyoto, Japan (
Tsuchiya. 2012), you could even make an argument that the combined overconsumption of beta carotene (mostly from supplements and enriched convenience foods) and the conditioned avoidance of the high fat foods that contain 'real' vitamin A may in fact be another maybe non-negligible contributer to the current obesity epidemic.
2.5x more all-trans-retinoic acid in chow reverse diet induced weight gainIn their manuscript that has been published online in advance, Tsuchiya et al. followed up on the results of a previous trial, in which genetically modified mice with almost no functioning vitamin A receptors showed a profound decrease in hepatic insulin-like growth factor-1 production and profound hepatic steatosis (fatty liver) - a pathology the scientists ascribed to profound insulin resistance as a direct consequence of the lack of vitamin A signaling. In their latest study, that was financed with a national research grant, and is soon to be published in the international journal
Hepatology (
Tsuchiya. 2012), Tsuchiya et al. fed C57BL/6J mice, which had been pre-fattened on the same high-fat, high-fructose diet many of our fellow human beings are indulging these days, diets containing either standard amount of vitamin A or 50mg of all-trans-retinoic acid per kg of chow.
 |
| Figure 1: Body weight (left) and glucose and insulin management (right) in normal and diabetes and obesity prone mice receiving control diet, standard high fat high fructose diet (HFHFr) or HFHFr + 50mg all-trans-retinoic acid (ATRA); data in the right is expressed relative to non-supplemented control (data calculated based on Tsuchiya. 2012) |
Not much to the researchers surprise, the administration of the high fructose high fat diet that contained 50mg/kg (normal chow has 20mg/kg), i.e. 2.5x more all-trans-retinoic acid, than the standard HFHFr chow, did not only stop the almost linear weight gain the animals had experienced in the course of the 16-week pre-fattening phase in normal mice (cf.
figure 1, left), it did also have statistically significant beneficial effects on the blood glucose (diabetes prone) and insulin (obesity prone) levels of mice that are genetically predisposed to develop diabetes (KK-Aγ) and obesity (ob/ob), in a second experiment.
The all-trans-retinoic acid (ATRA) <> leptin connectionAnd although the aforementioned results are certainly impressive, this is not an essentially novel finding - what was yet observed for the fist time in this study, is the reversal of the diet-induced reduction of hepatic leptin receptor expression in the HFHFr group receiving additional all-trans-retinoic acid in their diets (cf.
figure 2, left).
 |
| Figure 2: Effects of additional ATRA in diet on leptin receptor expression (left) and relative diet- and diet + supplementation induced changes in selected makers of non-alcoholic fatty liver disease (right; data adapted / calculated based on Tsuchiya. 2012) |
The latter went hand in hand with a restoration of IGF-1 BP2 and ameliorative effects on the measured markers of non-alcoholic fatty-liver disease markers (liver weight, lipid content; cf.
figure 2, right).
Vitamin(s) A - obesity and beyond These still "incompletely understood" (
Bonet. 2011) metabolic effects of 'real' vitamin A and its metabolites are yet only the tip of an iceberg of the largely ignored health effects of the first micronutrient in the vitamin alphabet. Other only partly related effects are
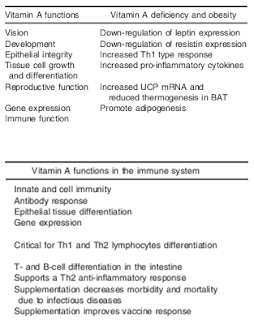 |
| Table 1: Vitamin A functions, major roles in the immune system and effects of vitamin A deficiency in undernutrition and obesity (Th1, T-helper type 1 response; Th2, T-helper type 2 response; UCP, uncoupling protein; BAT, brown adipose tissue; directly adapted from Garcia. 2012) |
- a coordinating / controlling effect of the enzyme that converts dietary vitamin A into the active form (ATRA) on lipid metabolism (Kiefer. 2012)
- a genetic blockade of adipocyte growth and thusly a direct inhibitory effect on die-induced obesity (Berry. 2012)
- a facilitative role in the maturation and replenishment of muscle progenitor cells (Ryan. 2011)
- an ability to prime pluripotent stem cells to become myocytes (muscle cells; Le May. 2011)
- anti-cancer effects (Streb. 2011; Siddikuzzaman. 2011)
- the repair of damaged heart muscle (Kikuchi. 2011; Freire. 2011)
- protective effects on cardio-myocytes against damage due to hyperglycemia (Guleria. 2011)
- systemic anti-inflammatory and immune regulatory effects via inhibition of interferon-gamma, TNF-alpha, NF-kappa-beta, IL-12, and promoting "an anti-inflammatory environment and adequate Th1:Th2 ratios" (Garcia. 2012)
- more general metabolic functions and immune-specific+ obesity-specific deficiency effects are summarized in table 1 (Garcia. 2012)
If we use the study at hand as a guide and the respective human equivalent dose of the ATRA contained in the 5g of chow/day the rodents consumed, the anti-NAFLD + anti-obesity effects would require a daily vitamin A intake of ~833IU for a mouse and 1,930IU /kg body weight for a human being, which is - I guess I don't have to tell you that - hilariously much and potentially hazardous.
How much vitamin A and where do you get it from?If we let ourselves be guided by the 2.5x amount of the standard dose (which was what the mice were actually fed), take the RDA as a reference for the latter and assume that the conversion of dietary vitamin A to all-trans-retinoic acid works properly, the corresponding human doses do actually seem pretty reasonable, with 11,250IU for men and 8,750IU for
non-pregnant women and can in fact be achieved relatively easily by eating, e.g.
- 25g chicken or pig liver or 30g of beef liver
- 100g butter + 300g cream + 300g cheddar cheese
- 200g of bluefin tuna + 3 eggs + tbsp of cod liver oil
In many cases it should thus suffice to simply forget your fat- and organ-meat phobia, to satisfy your retinol requirements - after all, the foods listed above may have the highest vitamin A content, but are by no means the only valuable sources of vitamin A
and reasonable amounts of its natural, plant-derived precursor beta carotene in a whole-foods diet.
 |
| Image 2: I don't know if you realize this, but all the good sources of vitamin D in this illustration also contain significant (measured in IU mostly way more) vitamin A. |
Unresolved issues with vitamin D: There was a time, when people used vitamin A to counteract vitamin D toxicity. Hard to imagine in the days of D-phoria, where everybody is advised to supplement, when a recent paper on the common measurement methods for vitamin D concluded that "several studies demonstrated that current 25(OH)D measurement methods do not meet" the prerequisite of being "sufficiently accurate over time, location and laboratory procedures" (
Tienpont. 2012). Recent research on the underlying physiological relation / antagonism of vitamin A and D is almost non-existent. The handful of rodent trials I am aware of that actually compared the effects of co-supplementation were conducted in the mid 20th century and yield no conclusive results as far as an 'optimal' A to D ratio would be concerned - mostly because they only elucidated how much vitamin A it would take to keep the rodents on vitamin D enriched diets alive for a few more days. A recently published study does yet suggest that the often heard recommendation to take 1,000IU of supplemental vitamin D3 per day does not effect vitamin A or leptin levels. Whether the -14% and -19% reductions in serum alpha- and gamma-tocopherol (vitamin E) levels that were observed in this trial (800IU D3 + 2g calcium) are physiologically significant would yet warrant further investigation (
Chai. 2012). The same is true for higher doses of vitamin D3, specifically, when those are - as it is often suggested taken in conjunction with those few fatty meals that actually contain 'real' vitamin A.
Assuming that you avoid supplementing high doses of beta carotene (don't care about the
various forms of carotenes in real food), as well as highly fortified convenience food (yeah, cereals belong to this category, as well, and may in fact be among the worst offenders) and do not fall for the idea that you need at least 10,000-20,000IU of supplemental vitamin D3 per day, simply because your skin is supposed to be capable of producing 10,000IU within less than 1h of sun exposure, you should get more then enough "raw material" for your body to produce ATRA from your diet.


