|
| Image 1: The word "inflammation" triggers associations which hinder a appropriate understanding of the complexities of the "inflammatory" immune response that is vitally important for (re-)building muscle tissue. |
Just to make sure that I do not get off another tangent, again, I will start right off, where I left you in the
last installment of the
Intermittent Thoughts and that was with the promise to have a closer look at the intricate relationship of (exercise-induced) inflammation and the increases in muscle-specific insulin-like growth factor 1 (IGF-1) and its splice variants, above all the muscle (re-)building mechano-growth factor 1 (MGF-1). Before we are looking how one influences the other,
we will yet have to establish a consistent understanding of "inflammation", which, despite being in on everyone's lips these days is commonly (mis-)understood and / or confused with "oxidation", as in the oxidation of "inflammable" substances, you have encountered innumerable times in the form of fire or rust.
What is inflammation? And is it good or bad?If we simply rely on our everyday understanding of inflammation, we are totally missing the boat on the true significance of a very complex net of biological processes some scientists quite blunderingly labeled "inflammation", which is not the "fire", i.e. the damaging (in many, but by no means all cases oxidative) process, itself,
but the appropriate, or, as in the case of auto-immune reactions, inappropriate physiological reaction to it. Whether this misleadingly termed reaction of your immune cells is "appropriate" and thusly healthy or "inappropriate" and thusly detrimental, depends on a whole host of factors, among which the distinction between
subclinical chronic inflammation and
acute inflammatory responses probably is the most important one.
 |
| Illustration 1: The theoretical relationship between the biphasic hormetic curve and exercise salience (Nunn. 2010. Fig. 1) |
While scientists believe that a chronic low, yet elevated level of inflammation is the root cause of almost all modern disease, the
acute inflammatory response to real threads is the driving force behind those hormetic adaptation processes about which Alistair V. Nunn and his colleagues from
Imperial College in London write that their "decline [...] in our daily life may be leading to increased systemic sub-clinical inflammatory tone, decreased metabolic flexibility and suppression of
exercise salience" and thusly set the stage for "obesity, the
metabolic syndrome, diabetes, vascular disease and even
cancer" (
Nunn. 2010). It is thusly only consistent of the researchers to demand:
Whether we like it or not, a long and healthy life needs to include regular exposure to occasional doses of environmental stressors, including fasting, natural temperature changes, polyphenols and exercise. Although human intelligence has enabled us to remove most stressors from the environment, common sense may be required to re-introduce some of them.
And while I could unquestionable go into much more detail on the concept of hormesis and its fundamental importance to our health, I am determined not to lose sight of the real intention of this installment of the
Intermittent Thoughts, which is to elucidate the intricate relationship between the
local inflammatory response to exercise, the intramuscular expression of IGF-1 and its splice variants and the exercise-induced increases in skeletal muscle mass and strength.
The IGF-1 response to acute inflammationContrary to what you may have gathered from a cursory read of the literature on the "dangers" of the "growth promoting" and thusly potentially carcinogenic insulin-like growth factor,
neither the mature 70 amino acid polypeptide IGF-1 nor any of its splice variants are in and out of themselves carcinogenic. It is the (not even indiscriminate, cf. red box) growth promoting effect they exert on target tissues via interactions with the respective IGF-1 receptors which will promote the growth and proliferation of all sorts of cells, including cancer cells that is responsible for their bad reputation.
 |
Image 2: IGF-1 per se is not fattening,
if anything it is "IGF-resistance" |
Did you know that a 2008 study by a group of scientists from the
University of Leipzig, in Germany, found that the "growth promoting" effect of IGF-1 on adipocytes is negligable, the effect of the latter on systemic IGF-1 expression via negative feedback, on the other hand pretty profound (
Klöting. 2008)? As it turned out,
not IGF-1, but its absence, or I should say, its inability to activate the receptor in the IGF-R knock-out mice that were used in the study were the underlying cause of both statistically significant increases in body, fat and organ weight, as well as ~20% elevated serum IGF-1 levels. Similar to the fattening effects of insulin, its
structural cousin (cf. insulin vs. insulin-
like growth factor discussion in the previous installment), it is thusly not the physiological expression of IGF-1, but its inability to trigger necessary cellular signaling cascades and negative feedback that could be at the heart of the metabolic derrangements that oftentimes go hand in hand with elevated levels of circulating IGF-1.
In this context an important result of a meta-study by Claudio Franceschi and his colleagueson genes involved in the etiology of longevity, comes to mind (
Franceschi. 2005):
In a longitudinal survey it has recently been shown that older women having low serum levels of IGF-I and high serum levels of IL-6 have the highest risk of disability and mortality, in comparison with women who have low levels of IL-6 and high levels of IGF-1 (Cappola et al., 2003). Such a beneficial effect of high IGF-1 serum level in the elderly is in apparent contrast with the above reported data showing that reduced IGF-I plasma levels are associated with longevity (Bonafè et al., 2003b). In order to reconcile this apparent discrepancy, it can be hypothesised that the decrease in plasma IGF-1 observed in nonagenarians and centenarians might minimise the risk of cancer in these subjects by decreasing a generalised mitogenic stimulation. The price to pay is frailty and massive reduction of muscle strength, two characteristics of such very old people.
With this connection between
overexpression of the inflammatory cytokine interleukine 6 (IL-6) and the low, or as we will see insufficient IGF-1 expression in elderly people, we have come full-circle and back to our initial question: How do "inflammation" and IGF-1 expression go together?
 |
| Image 3: Unlike Hermes, the Greek messenger of the Gods, cytokines have no intrinsically mischievous side and their vilification is unjust. |
Although it was certainly not a good idea to summarize such a complex phenomenon as the release of signaling molecules and the consequent reponse of the immune system under the term "inflammation",
the name "cytokine" is actually quite fitting, because the combination of the Greek words
-cyto, for "cell", and
-kinos, for "movement", denote the exact consequences the release of respective signaling molecules has: it induces the movement of cells, which, in the case of "inflammatory cytokines", obviously are immune cells. The
contemporary vilification of all "inflammatory" cytokines in the lay-press is however unwarranted - or would you hold the guy who takes the calls on the emergency line responsible for either the outbreak of the fire (=immune reaction necessary) or another nuisance alarm (unwanted auto-immune reaction)?
A very important clue that points us into the right direction comes from a 2007 study by Pelosi et al. (
Pelosi. 2007), who analyzed the regenerative process skeletal muscle tissue undergoes subsequent to injuries. The scientists analyzed the differential expression of the two major inflammatory cytokines
TNF-alpha and
IL-1-beta, which in turn triggers the release of the aforementioned (and much better known) IL-6 in skeletal muscle (
Luo. 2003), in response to
cartiotoxin (CTX) injection in normal (wild-type) mice and mice who were genetically engineered to over-express mIGF-1 specifically in differentiated myofibres (MLC/mIGF-1).
 |
| Figure 1: Differential expression (relative to maximum) of TNF-alpha and IL-1b in CTX-injected muscle of wild-type and MLC/IGF-1 mice during the 10 days of recovery (data adapted from Pelosi. 2007) |
As the data in figure 1 goes to show, the higher mIGF-1 expression (the "m-" indicates autocrine production, i.e. IGF-1 that is produced right at the target tissue, in this case skeletal muscle) in the genetically engineered mice led to a
statistically significant amelioration in the expression of pro-inflammatory cytokines, which are involved in the recruitment of monocytes and macrophages.
An "anomaly" you will probably have noticed is the sudden increase of both inflammatory marker on day 5 post injury. I don't know if you are familiar with the term "deep onset muscle soreness", but the "onset" increase in inflammation certainly reminds me of the feeling I tend to have whenever I have gone overboard on squatting. Do you know what I am talking about? This awkward feeling of cramping pain in the quads that tends to appear right then, when you thought that the soreness was abating? Interestingly enough, this sudden onset of inflammation, which is completely absent in the MLC/mIGF1 mice, goes hand in hand with a the peak of another, less well-known cytokine that goes by the (telling) name of macrophage migration inhibition factor, or MIF. This stands in contrast to the MIF response in the MLC/mIGF-1 mice, where
the significant down-regulation of MIF at 5 days post-CTX injection in MLC/mIGF-1 injured muscle may facilitate the emigration of infiltrating cell pools, leading to a rapid resolution of the inflammatory response.
These facilitatory, or rather dis-inhibiting effects IGF-1 seems to exert with respect to the MIF-driven "lockout" of the macrophages, allows for a "r
apid restoration of injured mIGF-1 transgenic muscle", of which Pelosi et al found that it...
was also associated with connective tissue remodeling and a rapid recovery of functional properties.
Show that autocrine mIGF1 via its modulating effect on the inflammatory response and its (related) ability to reduce the formation of fibrotic muscle tissue "creates a qualitatively different environment for sustaining more efficient muscle regeneration and repair" (
Pelosi. 2007).
 |
| Image 4: The local administration of platelet (and growth factor) rich plasma is about to become a recognized treatment strategy for muscular injuries and chronic degenerative joint diseases such as tendinopathy. |
Did you know that a 2006 study from the
University of Melbourne showed that both, IGF-1 gene transfer to the injured muscle (which would be comparable to the autocrine mIGF-1 expression discussed in the previous paragraph), as well as systemic IGF-1 administration via mini-osmotic pump at 1.5 mg/kg/day "hastened functional recovery" in artificially injured tibialis anterior muscles of mice? The injection of platelet rich plasma, which contains various growth factors, into injured muscle tissue is already practiced by many physicians working with competitive athletes (
Creany. 2007) and appears to be a promising treatment strategy for other (non-muscular) pathologies such as chronic degenerative tendinopathy, as well (
Vos. 2010).
If we set these results into a somewhat broader context, it becoms clear that the inflammatory cytokines that are released as a result of muscular damage, summon macrophages and other immune cells to the injured tissue. The concomitant production of
local mIGF-1 facilitates their migration into the muscle where they increase the proliferation of satellite cells (
Merly. 1999) and help (re-)building (new) muscle tissue (
Chazaud. 2003). The "ameliorative" effect of IGF-1 on inflammation is thusly by no means comparable to the "ameliorative" effect firefighters exert on a fire. IGF-1 does not work against the inflammatory response (remember: in 99% of all cases the latter is a completely healthy and beneficial physiological reaction to an external assault on your body!), it works hand in hand with the driving forces of "inflammation", the monocytes, by "opening the door to the muscle" and rejuvenating the satellite cell pool from which, in turn, relies on the immune cells during the incorporation of these progenitor cells into the existing muscle tissue.
The emerging importance of an endocrine-immune-axis in skeletal muscle hypertrophy  |
| Image 5: Control (A) and IL-15 treated (B) myotubes; nuclei are stained yellow; note the wide myotubes in the IL-15 treated muscle (img. from Quinn. 2002) |
This intricate interplay of the endocrine (IGF) and the immune (monocytes) system, which is so characteristic for our emerging understand of the true complexity of the mammalian physiology, reminds me of the question Trevor's Facebook question from last week. Trevor, who has obviously done his homework on the "IGF-1 / cytokine connection" wanted to know my thoughts on interleukin-15, one of the less-researched "inflammatory" cytokines, which appears to play a central role in the accrual of myosin heavy chain (MHC) motor proteins (if you have not done so, already you can read more about the role of the motor proteins in
Part II of the Hypertrophy 101). Back in 1995, already, a group of scientists from the
American Lake VA Medical Center published a ground-breaking (yet hitherto unfortunately largely overlooked) paper on the role of interleukin-15 in skeletal muscle myogenesis (
Quinn. 1995). Quinn et al. were for the first time able to show that
IL-15 used at concentrations of 10 or 100 ng/ml increased MHC accumulation five-fold in C2 myoblast cultures and 2.5-fold in primary bovine myogenic cultures. Moreover, C2 myotubes formed in the presence of IL-15 appeared larger than controls.
Interestingly, the researchers must have apprehended the existence of the previously discussed intreaction of the endocrine and the immune system and tested whether this effect depended on the presence of IGF-1:
 |
| Figure 2: Moysin heavy chain expression (arbitrary units) in in bovine muscle cultures after incubation with IL-15 (dose in ng/ml), IGF-1 (dose in ng/ml) or both (data adapted from Quinn. 1995). |
From the data in figure 2 it becomes quite obvious that IL-15 has more than a facilitative effect on the IGF-1 induced accrual of motor proteins. A 2002 follow up study on mice myocytes (
Quinn. 2002) and a 2003 study using human skeletal muscle myogenic cultures (
Quinn. 2003) confirmed the validity of these initial findings.
 |
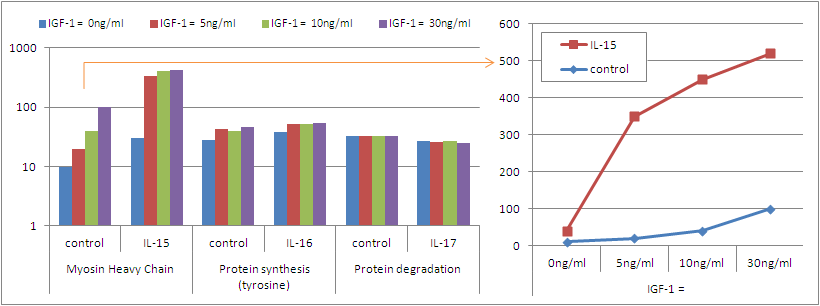
| Figure 3: Myosin heavy chain expression, protein synthesis and protein degradation in rodent muscle in response to IL-15 treatment at different basal levels of IGF-1 (data adapted from Quinn. 2002) |
Interestingly, the synergistic effect of IL-15 and IGF-1 appears to be restricted to the accrual of motor proteins (cf. figure 3) and has
only marginal effects on protein synthesis and degradation.
mTOR & Co, IGF-1, inflammation ... what's next? |
| Image 6: Is the role of naturally achievable testosterone levels in the accrual of lean muscle tissue overrated, or not? What exactly does the principal male androgen do on a tissue level and why did your OTC test booster only increase your libido and not the size of your sleeves? Come back on 01.01.2012 to learn more ;-) |
With protein synthesis and degradation, we have come back to one of the initial discussed cornerstones of skeletal muscle hypertrophy (cf.
What is Hypertrophy?), of which you should have learned in the previous installment of this series that is
a necessary, yet not sufficient prerequisite of sustainable muscle growth. Without the IGF-1 mediated and, as you have learned in this installment, monocyte-driven (re-)construction (increase in myonuclei + accumulation of motor proteins) of the underlying structure of the muscle, however, neither the repair of damaged, nor the accrual new,
functional (cf.
Hypertophy 101: Part II) muscle tissue would be possible.
The question we still have to answer before we can eventually integrate all those different pathways into a model which would allow us to develop a "hypertrophy-optimized" training, nutrition and supplementation regimen,
we do yet still have to shed some light on the role of the legendary "big T": Testosterone! So stick with me and
come back next week, or next year, whatever you like better, to learn more about the actual role of the principal male sex in the complex process of skeletal muscle growth.